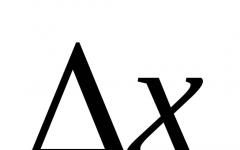Образовательный портал. Схема наследования окраски пера у кур Наследование окраски оперения у кур
Полное доминирование
Полное доминирование проявляется когда доминантная аллель полностью скрывает присутствие рецессивной аллели (1 закон Менделя), т. е. в генотипе гетерозигот присутствует продукт одного гена.
Неполное доминирование
Неполное доминирование проявляется когда в фенотипе гетерозигот (Аа) появляется промежуточный (средний) признак. Расщепление по фенотипу и генотипу при неполном доминировании одинаково соответствует (1АА:2Аа:1аа – по генотипу, соответствует 1красный:2 розовый:1 белый – по фенотипу).
Рассмотрим пример неполного доминирования окраски цветков ночной красавицы:
Фенотипы Р: красные цветки х белые цветки
Генотипы Р: АА х аа
Генотип F1: Аа
Фенотип F1: розовые цветки
Генотипы F1: Аа х Аа
ФенотипыF1: розовый х розовый
G: А, а А, а
Расщепление F2 по фенотипу -1красный:2 розовый:1 белый, по генотипу -1АА:2Аа:1аа
Множественный аллелизм
Явление существования более двух альтернативных аллельных генов, имеющих различные проявления в фенотипе. Кроме основных – доминантного и рецессивного - генов, появляются промежуточные, которые по отношению к доминантным ведут себя как рецессивные, а по отношению к рецессивным – как доминантные.
В приведенных выше закономерностях наследования один ген представлен двумя аллелями (доминантный А и рецессивный – а), определяет проявление одного признака (классические Менделеевские). Эти состояния гена возникают вследствие мутирования. Ген А может мутировать неоднократно, и возникает целая серия аллельных генов (а 1 , а 2 , а 3 ….а n) – множественный аллелизм, что характеризует разнообразие генофонда целого вида (совокупности всех генов в составе генотипов всех особей вида). А в генотипе отдельного диплоидного организма могут находиться только 2 гена из серии аллелей (Аа 1 , Аа 2 , Аа, аа 1 и т.д.). Множественный аллелизм является видовым, а не индивидуальным признаком (в отличие от полимерии).
Т.Л. Богданова предлагает рассмотреть пример множественного аллелизма на примере трехчленной серии аллелей окраски шерсти у кроликов:
1-Сплошная темная окраска.
2 - Белая (альбинизм –полное отсутствие пигментации шерсти).
3 - Горностаевая (гималайская) –общая белая окраска тела, а кончики ушей, лап и мордочка – черные.
Ген сплошной окраски доминирует над остальными членами серии, ген горностаевой окраски доминантен по отношению к белой, но рецессивен по отношению к сплошной темной. Ген белой окраски рецессивен по отношению и к сплошной, и к горностаевой (1> 2> 3).
А.А. Кириленко предлагает рассмотреть пример множественного аллелизма на примере окраски шерсти у кроликов четырехчленной серии:
Черные – ген А, белые – ген аа (гомозиготы рецессивные). Но существуют еще несколько состояний этого гена, имеющие собственный фенотип в гомозиготе – шиншиловый – сплошная серая окраска и гималайский (горностаевый) – белый, кончики ушей, хвоста, лап и носа – черные. В гомозиготе шиншиловый – а ch а ch , а гиммалайский а h а h .
А > а ch > а h > а
Кодиминирование
Кодоминированием называют такое взаимодействие аллельных генов, когда у гетерозигот в фенотипе присутствует продукт обоих генов. Гены одной аллельной пары равнозначны, ни один ни другой не подавляет действие другого, если оба находятся в генотипе, то оба проявляют свое действие.
Примером кодоминирования является наследование группы крови у человека. Группа крови у человека контролируется аутомомными генами I и i и имеет три аллели – О, А и В. Аллели А и В являются доминантными, а аллель О (ноль) рецессивна по отношению к обеим доминантным аллелям.
Каждый доминантный ген контролирует синтез в эритроцитах соответствующих белков-агглютиногенов А или В (при О агглютиногены не синтезируются). При наличии у человека двух доминантных аллелей А и В имеет место кодоминирование и в эритроцитах синтезируются оба белка-агглютиногена. Таким образом, группа крови определяется наличием или отсутствием того или иного белка-агглютиногена.В
| Генотипы | Фенотипы | Генотипы | ||||
| Аллельные гены | i o | I A | I B | I Группа (О) | i o i o | ОО |
| I o | i o i o | I A i o | I B i o | II Группа (А) | I A i o , I A I A | АО, АА |
| I A | I A i o | I A I A | I A I B | III Группа (В) | I B i o , I B I B | ВО, ВВ |
| I B | I B i o | I A I B | I B I B | IV Группа (АВ) | I A I B | АВ |
Сверхдоминирование
Сверхдоминирование наблюдается если доминантный ген в гетерозиготном состоянии проявляет свое действие сильнее, чем в гомозиготном.
Пример сверхдоминирования: у дрозофилы есть летальный ген а (гомозиготы аа погибают). Гомозиготы по гену А (АА) имеют нормальную жизнеспособность, а гетерозиготы (Аа) живут дольше и более плодовиты, чем доминантные гомозиготы.
Комлемнтарность
Взаимодействие нескольких неаллельных генов, приводящее к развитию нового признака, отсутствующего у родителей. Неаллельные гены в этом случае как бы дополняют друг друга, в результате чего развивается совершенно иной признак. Комплементарными называют гены, обуславливающие при совместном сочетании в генотипе в гомозиготном или гетерозиготном состоянии новые фенотипические признаки. Особенности расщепления потомства от скрещивания при комплементарном взаимодействии генов зависит от того, имеют ли эти гены самостоятельное фенотипическое проявление (далее примеры приводятся по О.В.Гончарову) :
- 9: 3: 3: 1 – каждый доминантный ген имеет самостоятельное фенотипическое проявление, сочетание в генотипе двух этих генов обуславливает развитие нового фенотипического проявления, а их отсутствие – не дает развитие признака. Пример: наследование формы гребня у кур. У домашних кур возможны четыре формы гребня: розовидный, гороховидный, ореховидных и простой (листовидный). Развитие этого признака связано с взаимодействием двух пар неаллельных генов. Розовидный гребень образуется при наличии доминантного гена одной аллели – А, гороховидный – действием доминантного гена другой аллели – В. У гибридов при наличии двух доминантных неаллельных генов АВ развивается ореховидный гребень, а при отсутствии всех доминантных генов, т. е. рецессивных гомозигот, по двум неаллельным генам – ааbb – образуется простой гребень.
Фенотипы Р: розовидный гребень х гороховидный гребень
Генотипы Р: ААbb x aaBB
Генотип F 1: AaBb
Фенотип F 1: ореховидный гребень (единообразие F 1 , появляется новый признак)
Фенотипы F 1: ореховидный гребень х ореховидный гребень
Генотипы F 1: AaBb x AaBb
| Гаметы | AB | Ab | aB | ab |
| AB | AABB ореховидный | AABb ореховидный | AaBB ореховидный | AaBb ореховидный |
| Ab | AABb Ореховидный | AAbb розовидный | AaBb ореховидный | Aabb розовидный |
| aB | AaBB Ореховидный | AaBb ореховидный | aaBB гороховидный | aaBb гороховидный |
| ab | AaBb ореховидный | Aabb розовидный | aaBb гороховидный | аabb простой |
Происходит расщепление на четыре фенотипические группы – в соответствии с фенотипическим радикалом дигибридного скрещивания:
9 A_B_ : 3 A_bb: 3 aaB_ : 1 aabb
ореховидный розовидный гороховидный простой
- 9: 7 – доминантные и рецессивные аллели комплементарных генов не имеют самостоятельного фенотипического проявления. Например, пурпурная окраска венчика цветка у душистого горошка развивается только при совместном сочетании в генотипе доминантных генов A и B , во всех остальных случаях окраска отсутствует и венчик оказывается белым.
Фенотипы Р: белый цветок х белый цветок
Генотипы Р: AAbb x aaBB
Фенотип F 1: пурпурный цветок
Генотип F 1: AaBb
F 1 (фенотипы): пурпурный х пурпурный
F 1 (генотипы): AaBb x AaBb
G: AB, Ab, aB, ab AB, Ab, aB, ab
Происходит расщепление на две фенотипические группы:
9 (A_B_) : 7 (aaB_, A_bb, aabb)
Пурпурный цветки Белые цветки
- 9: 3: 4 – доминантные и рецессивные аллели комплементарных генов имеют самостоятельные фенотипические проявления. Например окраска зерен у ржи: родители имеют окраску зерен: белую и желтую. Гибриды первого поколения – AaBb- имеют зеленый цвет зерен (новый признак). Гибриды первого поколения скрестили между собой и получили во втором поколении расщепление по окраске зерен: 9 зеленых:3 желтых: 4белых.
Зерно белого цвета – aaBB, aabb , зерно желтого цвета- AAbb , зерно зеленого цвета – AABB,AaBb
Фенотип F 1: зеленая окраска зерен
Генотип F 1: AaBb
F 1 (фенотипы): зеленая х зеленая
F 1 (генотипы): AaBb x AaBb
G: AB, Ab, aB, ab AB, Ab, aB, ab
Происходит расщепление на четыре фенотипические группы:
9 A_B_ : 3 A_bb: 4 (aaB_,aabb)
зеленый желтый белый
- 9: 6: 1 – сочетание доминантных аллелей комплементарных генов обеспечивает формирование одного признака, сочетание рецессивных аллелей этих генов другого, а сочетание в генотипе только одного из доминантных генов – третьего признака. Например, у тыквы встречаются три типа плода: дисковидную, удлиненную (грушевидную), округлую (сферическую) . Родители имеют округлую форму плодов – Aabb и aaBB. Гибриды первого поколения имеют дисковидные плоды. При скрещивании гибридов первого поколения образуются следующие сочетания: 9 с генотипом A_B_ имеют дисковидную форму плода, 1 с генотипом aabb – удлиненную (грушевидную) форму плода, а 6 с генотипами A_bb и aaB_ - имеют округлую (сферическую) форму плода. Форма тыквы контролируется двумя парами неаллельных генов. Доминантные аллели каждой из пар отдельно определяют развитие округлой формы (А и В), а их рецессивные аллели – развитие грушевидной формы (a и b). Сочетание двух доминантных неаллельных генов приводит к развитию дисковидной формы тыквы (АВ).
Фенотипы Р: округлая форма х округлая форма
Генотипы Р: AAbb x aaBB
Фенотип F 1: дисковидная форма
Генотип F 1: AaBb
F 1 (фенотипы): дисковидная х дисковидная
F 1 (генотипы): AaBb x AaBb
G: AB, Ab, aB, ab AB, Ab, aB, ab
Происходит расщепление на три фенотипические группы:
9 (A_B_) : 6 (aaB_, A_bb) : 1 (aabb)
дисковидная форма округлая форма грушевидная форма
А.А. Кириленко разделяет «комплементарность» на «кооперацию» (пример с наследованием формы гребня у кур –9:3:3:1) и «комплементарное действие генов» (пример с наследованием окраски венчика у душистого горошка -9:7; пример с наследованием формы плодов тыквы – 9:6:1; пример наследования окраски зерен у ржи -9:3:4)
Эпистаз
Эпистатичным называют такое влияние неаллельных генов, при котором один из них подавляет действие двугого. Ген, подавляющий действие другого, неаллельного гена, называют супрессором (ингибитором) и обозначается буквами I или S. Подавляемый ген называют гипостатичным. Супрессор может быть доминантным (доминантный эпистаз) и рецессивный (криптомерия).
Эпистатичное взаимодействие связано с синтезом регуляторных белков. В одном случае регуляторный ген синтезирует белок, который подавляет процесс транскрипции основного структурного гена, ответственного за развитие признака. В другом случае регуляторный ген, наоборот, не способен синтезировать белок, который регулирует активность структурного гена. И в том и в другом случае признак развивается, так как белок, ответственный за признак, не синтезируется.
Доминантным эпистазом называют подавление действия гена доминантной аллелью другого гена (доминатный ингибиторный ген I > А). Расщепление потомков F 2 при доминантном эпистазе может быть различным:
- 13:3 – расщепление наблюдается в том случае, если доминантная аллель эпистатичного гена не имеет своего фенотипического проявления, а лишь подавляет действие другого гена, в то время когда рецессивная аллель не влияет на проявление признака. Пример: окраска кур (ген А отвечает за черную окраску, ген I подавляет действие гена А).
Р (фенотипы): курица белая х петух белый
Р (генотип): ААII x aaii
F 1: AaIi - белая окраска кур
F 1 (генотипы): курица AaIi х петух AaIi
G: AI, Ai, aI, ai AI, Ai, aI, ai
- 12:3:1 – расщепление наблюдается в том случае, если гомозиготная по рецессивным признакам особь имеет особый фенотип (самостоятельное фенотипическое проявление). Например, доминантный эпистаз наблюдается при наследовании окраски шерсти у спаниелей. У собак окраска шерсти может быть черной (доминантный признак - A) и коричневой (рецессивный признак - a). Развитие любой окраски ингибирует (подавляет) доминантный ген – I, в этом случае шерсть будет иметь белую окраску (пигмент отсутствует). Его рецессивный аллель – i никак не влияет на развитие признака. Рассмотрим результаты скрещивания коричневой самки с белым самцом. В F1 все щенки были белыми, а при скрещивании гибридов первого поколения появились 12 белых, 3 черных и 1 коричневый щенок. Появление черных особей свидетельствует о наличии в генотипе белой собаки генов доминантной окраски, которые в фенотипе не проявляются.
Р (фенотип): самка коричневая х самец белый
Р (генотип): iiaa x IIAA
F1: IiAa – белая окраска шерсти
F1(генотипы): самка IiAa х самец IiAa
G: IA , Ia , iA, ia IA , Ia , iA, ia
Рецессивным эпистазом называют такое взаимодействие неаллельных генов, при котором рецессивная аллель эпистатичного гена в гомозиготном состоянии подавляет действие другого гена (рецессивный ингибиторный ген ii > А).
При одинарном рецессивном эпистазе рецессивная аллель одного гена подавляет действие другого (аа подавляет В_)
При двойном рецессивном эпистазе рецессивная аллель каждого гена в гомозиготном состоянии подавляет действие доминантной аллели другого (аа подавляет В_, bb подавляет А_).
При рецессивном эпистазе в потомстве наблюдается расщепление 9:3:4 или 9:7 (такое же расщепление характерно и для комплементарного взаимодействия некоторых генов – определить характер наследования признаков в случае рецессивного эпистаза можно только сочетая гибридологический анализ с изучением биохимии и физиологии развития данного признака).
Рецессивным эпистазом объясняется «бомбейский феномен» - необычное наследование антигенов системы групп крови АВО. В семье женщины с I группой крови (I o I o) и от мужчины со II группой крови (I A I A) родился ребенок, имеющий IVгруппу крови (I A I B), а это невозможно. Оказалось, что женщина унаследовала от своей матери ген I B , а от своего отца ген I O и у нее должна быть III группа крови. Но проявил действие только ген I O , а ген I B был подавлен рецессивным геном х, который находился в гомозиготном состоянии – хх. Этим можно объяснить то, что у женщины фенотипически проявилась I группа крови. А у ребенка этой женщины подавленный ген I B проявил свое действие. Ребенок имел IV группу крови I A I B .
Р (бабушки и дедушки): I B I B Xx (IIIгруппа) x I O I O Xx (I группа)
G: I B X I B x I O X I O x
F1: I B I O xx (I группа)
F1: женщина I B I O xx (Iгр) x мужчина I A I A XX (IIгр)
G: I B x I O x I A X
F2: I A I B Xx (IV группа)
Другим примером рецессивного эпистаза является наследование окраски шерсти у мышей (по А.А. Кириленко, по Т.А. Шустановой это пример комплементарного взаимодействия). Родители: черная мышь и белая мышь. Все первое поколение – серые – CcIi. Самцов и самочек первого поколения скрестили между собой и получили по втором поколении расщепление 9 серых: 3 черных: 4 белых.
Черная окраска – ссI_; серая окраска (агути) – С_I _; белая окраска – С_ ii или ссii
F1: самочка CcIi x самец CcIi
G: CI, Ci, cI, ci CI, Ci, cI, ci
Полимерия
Это явление, когда несколько (два и более) неаллельных генов –- отвечают за развитие одного и того же признака, причем степень развития признака зависит от общего количества доминантных генов (чем больше одинаковых аллелей, тем интенсивней проявляется признак. Эти гены называют полимерными (множественными) и обозначают одинаковыми буквами с соответствующим индексом (например, А 1 ,А 2). Чаще всего полимерные гены контролируют количественные признаки (высота растений, масса животных, яйценоскость кур, длина волокна у льна, содержание сахара в корнях сахарной свеклы и др.), но бывают и качественные (цвет кожи у человека).
Полимерия может быть кумулятивной (аддитивной, суммирующей) и некумулятивной.
Кумулятивная полимерия при ней степень проявления признака зависит от числа доминантных аллелей соответствующих полимерных генов, содержащихся в генотипе особи (чем больше доминантных генов, тем выше степень проявления признака) – расщепление в потомстве наблюдается 1:4:6:4:7. Примеры: наследование цвета кожи у человека, яйценоскости кур, наследование окраски зерна разных сортов пшеницы.
У негроидов в двух парах генов все четыре аллели доминантные (А 1 А 1 А 2 А 2), а у европеоидов с белой кожей все аллели рецессивные (а 1 а 1 а 2 а 2). Сочетание разного количества доминантных аллелей приводит к образованию мулатов с разной интенсивностью окраски кожи – от темной до светлой.
Фенотипы (Р): европеоид х негроид
G: а 1 а 2 А 1 А 2
Генотип F 1: А 1 а 1 А 2 а 2
Фенотип F 1: средний мулат
F 1 (фенотип): средний мулат средний мулат
Во втором поколении наблюдается расщепление на пять фенотипических групп:
1 негр: 4 темных мулата: 6 средних мулатов: 4 светлых мулата: 1 белокожий
Пример наследования окраски зерна пшеницы: эксперимент проводил шв. генетик и селекционер Герман Нильссон-Эле – скрещивал разные сорта пшеницы с красным и белым зерном. Во втором поколении он чаще всего обнаруживал расщепление в соотношении 3:1, но в некоторых случаях (если сорта отличались по трем парам аллелей) - растений с белым зерном оказывалось значительно меньше (расщепление шло 15 красных:1белое). Отличались между собой, интенсивностью окраски, и красные зерна (от красной до светло-розовой), так как растения имели в генотипе разное количество доминантных генов, отвечающих за красную окраску. Растения с белым зерном в генотипе имели только рецессивные гены. Поэтому данный пример у разных авторов вы можете увидеть как пример кумулятивной полимерии (1:4:6:4:1), а у других авторов как пример некумулятивной полимерии (15:1).
Фенотипы Р: красное зерно х белое зерно
Генотипы Р: А 1 А 1 А 2 А 2 А 3 А 3 х а 1 а 1 а 2 а 2 а 3 а 3
G: А 1 А 2 А 3 а 1 а 2 а 3
F1 (фенотип): 100% розовое зерно
F1 (генотип): А 1 а 1 А 2 а 2 А 3 а 3
F2 (фенотип):
1 вариант расшифровки результатов используется как кумулятивная полимерия :
2 вариант расшифровки результатов используется как пример некомулятивной полимерии :
Некумулятивная полимерия – степень развития признака зависит не от количества доминантных генов, а лишь от их наличия в генотипе. Пример: куры с генотипом а 1 а 1 а 2 а 2 – имеют неоперенные ноги, во всех остальных случаях – ноги оперены, причем степень оперенности ног одинаковая при любом числе доминантных генов (расщепление в потомстве наблюдается 15:1); наследование формы стручков у пастушьей сумки (качественный признак) -15:1 и наследование глухоты у человека – 15:1.
Пример наследования у кур признака «оперенные ноги»
Фенотипы (Р): неоперенные ноги х оперенные ноги
Генотипы (Р): а 1 а 1 а 2 а 2 х А 1 А 1 А 2 А 2
G: а 1 а 2 А 1 А 2
Генотип F 1: А 1 а 1 А 2 а 2
Фенотип F 1: оперенные ноги
F 1 (генотипы): А 1 а 1 А 2 а 2 х А 1 а 1 А 2 а 2
F 1 (фенотип): оперенные ноги оперенные ноги
G: А 1 А 2 , А 1 а 2 , а 1 А 2 , а 1 а 2 А 1 А 2 , А 1 а 2 , а 1 А 2 , а 1 а 2
| гаметы | А 1 А 2 | А 1 а 2 | а 1 А 2 | а 1 а 2 |
| А 1 А 2 | А 1 А 1 А 2 А 2 Оперенные | А 1 А 1 А 2 а 2 оперенные | А 1 а 1 А 2 А 2 оперенные | А 1 а 1 А 2 а 2 оперенные |
| А 1 а 2 | А 1 А 1 А 2 а 2 Оперенные | А 1 А 1 а 2 а 2 оперенные | А 1 а 1 А 2 а 2 оперенные | А 1 а 1 а 2 а 2 оперенные |
| а 1 А 2 | А 1 а 1 А 2 А 2 Оперенные | А 1 а 1 А 2 а 2 Оперенные | а 1 а 1 А 2 А 2 оперенные | а 1 а 1 А 2 а 2 оперенные |
| а 1 а 2 | А 1 а 1 А 2 а 2 оперенные | А 1 а 1 а 2 а 2 оперенные | а 1 а 1 А 2 а 2 оперенные | а 1 а 1 а 2 а 2 неоперенные |
Во втором поколении наблюдается расщепление на две фенотипические группы:
15 с оперенными ногами: 1 с неоперенными ногами
Плейотропия
множественное действие гена – явление при котором один ген одновременно влияет на формирование нескольких признаков.
В рассмотренных ранее вариантах скрещивания (классических менделеевских) вы познакомились с однозначным действием гена, которое реализуется по схеме:
Если один ген оказывает влияние на развитие нескольких признаков, говорят о множественном действии гена, которое идет по другой схеме:
Механизм множественного действия гена заключается в том, что белок, кодируемый этим геном, может в большей или меньшей степени участвовать в различных, мало связанных между собой процессах жизнедеятельности.
Впервые на это явление обратил внимание Г. Мендель. Впоследствии плейотропия была обнаружена у многих организмов.
| Организм | Наличие гена | Влияние гена на другие признаки |
| У человека | Ген, отвечающий за рыжую окраску волос | - отвечает за отсутствие пигмента меланина в коже (светлая кожа); - отвечает за появление веснушек. |
| У человека | Ген, вызывающий наследственное заболевание - синдром Морфана | - нарушает развитие соединительной ткани; - удлиняются фаланги пальцев; - возникают аномалии в строении хрусталика; - наблюдаются нарушения в сердечно-сосудистой системе. |
| У дрозофилы | Ген белой окраски глаз | - влияем на цвет тела и внутренних органов; - влияет на длину крыльев; - влияет на строение полового аппарата; -снижает плодовитость; - уменьшает продолжительность жизни. |
| У свеклы | Ген, отвечающий за окраску корнеплода | - влияет на цвет черешка листьев; - влияет на цвет жилок листа |
При множественном действии одного гена могут иметь место два варианта расщепления двух признаков. В одном случае оба признака могут проявляться в соотношении 3 (АА, Аа, Аа) : 1 (аа). В другом случае один признак проявляется в соотношении 3 (АА, Аа, Аа) : 1 (аа), а другой признак в соотношении 1(АА) : 3 (Аа,Аа,аа).
| I В А Р И А Н Т | I В А Р И А Н Т | ||||||
| 1 признак | 1 признак | ||||||
| АА | Аа | Аа | аа | АА | Аа | Аа | аа |
| 2 признак | 2 признак | ||||||
Это связано с тем, что гены, определяющие морфологические признаки, влияют на физиологические функции (изменяют жизнестойкость, плодовитость особей) или оказываются летальными (вызывают гибель зародыша или организма при его рождении). Например, ген, определяющий голубую окраску у норки, снижает ее плодовитость. Доминантный ген серой окраски у каракулевых овец в гомозиготном состоянии летален, так ягнята рождаются с недоразвитыми желудками и погибают при переходе на грубые корма (травой).
Множественное действие генов
Как известно, доминирование есть подавление действия одной аллели другой аллелью, представляющих собой один ген: А > а, В > b, С > с и т. д.
Но существует взаимодействие, при котором аллель одного из генов подавляет действие аллелей других генов, например А > В или В А, а > В или b > А и т. д. Такое явление «доминирования» между генами называется эпистазом .
Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному взаимодействию.
Гены, подавляющие действие других генов, называются супрессорами, или ингибиторами. Они могут быть как доминантными, так и рецессивными. Гены-супрессоры известны у животных (млекопитающие, птицы, насекомые) и у растений. Обычно они обозначаются I или Su в случае доминантного состояния генов и i или su для их рецессивных аллелей (от английских слов inhibitor или supressor).
В настоящее время эпистаз делят на два типа: доминантный и рецессивный.
Под доминантным эпистазом понимают подавление доминантной аллелью одного гена действия аллельной пары другого гена. Из многих примеров доминантного эпистаза, установленных как на животных, так и на растениях, приведем лишь отдельные.
Некоторые породы кур имеют белое оперение (белый леггорн, белый примутрок и др.), другие же породы имеют окрашенное оперение (австралорп, ньюгемпшир, полосатый плимутрок и др.). Белое оперение разных пород кур определяется несколькими различными генами. Так, например, доминантная белая окраска определяется генами CCII (белые леггорны), а рецессивная белая - ccii (белые суссексы, белые минорки, белые плимутроки). Ген С определяет наличие предшественник пигменту (хромогена), т. е. окрашенность пера, его аллель с - отсутствие хромогена и, следовательно, неокрашенность пера птицы. Ген I является подавителем действия гена С, аллель i не подавляет его действия. В присутствии даже одной дозы гена I в генотипе птицы действие генов окраски не проявится. Поэтому при скрещивании белых леггорнов CCII с цветными породами CCii, как правило, доминирует белая окраска CCIi. При скрещивании белых плимутроков ccii с окрашенными породами CCii гибриды оказываются окрашенными Ccii. Следовательно, у леггорнов белая окраска является доминантной, а у плимутроков - рецессивной.
Если же производится скрещивание белых леггорнов CCII с рецессивными белыми плимутроками ccii, то в первом поколении цыплята оказываются тоже белыми CcIi. При скрещивании между Собой гибридов F 1 во втором поколении имеет место расщепление по окраске в отношении 13 / 16 белых: 3 / 16 окрашенных.
Как можно объяснить полученное отношение? Прежде всего, данное отношение говорит о расщеплении по двум генам, это расщепление можно представить как 9(С-I-) + 3(ccI-) + 1(ccii) = 13 и 3(С-ii), что соответствует формуле 9:3:3:1.
Очевидно, в этом случае окраска леггорнов обязана не присутствию особых генов белой окраски, а действию гена - подавителя окраски (I-). Тогда генотип гомозиготных белых леггорнов должен быть ССII, где I является геном - подавителем окраски, а С - геном окраски. Белые плимутроки по генотипу должны быть гомозиготными по двум рецессивным факторам ccii, где с - отсутствие окраски и i - отсутствие подавления окраски. В силу эпистатирования I > С гибридные куры первого поколения CcIi должны быть белыми. В F 2 все куры с генотипами 9 / 16 С-I-, 3 / 1 6 ccI- и 1 / 1 6 ccii также должны быть белыми, и лишь куры одного фенотипического класса 3 / 1 6 (С-ii) оказываются окрашенными, поскольку здесь содержится ген окраски и нет его подавителя.
Таким образом, подавление действия доминантной аллели гена, Определяющего развитие окраски, доминантной аллелью другого гена (подавителем) обусловливает в F 2 расщепление по фенотипу в отношении 13: 3.
Доминантный эпистаз может давать и другое отношение при расщеплении в F 2 по фенотипу, а именно 12: 3: 1. В этом случае форма гомозиготная по обоим рецессивным факторам aabb, будет фенотипически отличима от форм с доминантными аллелями двух генов А-В- и форм с одной из них: ааВ- и А-bb. Такое расцепление установлено для наследования окраски плодов у тыквы, кожуры у лука и других признаков. В этом случае в расщеплении также принимает участие доминантный ингибитор.
Мы разобрали взаимодействие только двух генов. В действительности по типу эпистаза взаимодействуют многие гены. Гены-подавители обычно не определяют сами какой-либо качественной реакции или синтетического процесса, а лишь подавляют действие других генов. Однако когда мы говорим, что ген-подавитель не имеет своего качественного влияния на признак, то это относится только к данному признаку. На самом же деле ингибитор, подавляя, например, пигментообразование, может оказывать плейотропное действие на другие свойства и признаки.
Под рецессивным эпистазом понимают такой тип взаимодействия, когда рецессивная аллель одного гена, будучи в гомозиготном состоянии, не дает возможности проявиться доминантной или рецессивной аллелям других генов: аа > В или aa > bb.
Мы уже имели случай познакомиться с расщеплением 9:3:4 как результатом . Но эти же случаи можно рассматривать и как пример рецессивного эпистаза.
При скрещивании черных мышей (ААbb) с белыми (ааВВ) все особи F 1 (АаВb) имеют окраску типа агути, а в F 2 9 / 16 всех особей сказываются агути (А-В-), 3 / 16 черные (A-bb) и 4 / 16 белые (ааВ- и aabb). Эти результаты можно объяснить, предположив, что имеет место рецессивный эпистаз типа ааВ-. При этом мыши генотипа ааВ- оказываются белыми потому, что ген а в гомозиготном состоянии, обусловливая отсутствие пигмента, препятствует тем самым проявлению гена-распределителя пигмента В.
Кроме описанных случаев одинарного рецессивного эпистаза, действуют и такие, когда рецессивная аллель каждого из генов в гомозиготном состоянии одновременно реципрокно подавляет действием доминантных аллелей каждого из генов, т. е. аа эпистатирует над В-, a bb над А-. Такое взаимодействие двух подавляющих рецессивных генов называют двойным рецессивным эпистазом . При этом в дигибридном скрещивании расщепление по фенотипу будет соответствовать 9: 7, как и в случае комплементарного взаимодействия генов.
Следовательно, одно и то же отношение можно трактовать и как комплементарное взаимодействие, и как эпистатирование. Сам по себе генетический анализ наследования при взаимодействии генов без учета биохимии и физиологии развития признака в онтогенезе не может раскрыть природы этого взаимодействия. Но без генетического анализа нельзя понять наследственной детерминации развития этих признаков.
Теперь обратимся к проблеме взаимодействия неаллельных генов. Если развитие признака контролируется более чем одной парой генов, то это означает, что он находится под полигенным контролем. Установлено несколько основных типов взаимодействия генов: комплементарность, эпистаз, полимерия и плейотропия.
Первый случай неаллельного взаимодействия был описан в качестве примера отклонения от законов Менделя английскими учеными У. Бетсоном и Р. Пеннетом в 1904 г. при изучении наследования формы гребня у кур. Различные породы кур характеризуются разной формой гребня. Виандотты имеют низкий, правильный, покрытый сосочками гребень, известный под названием “розовидного”. Брамы и некоторые бойцовые куры обладают узким и высоким гребнем с тремя продольными возвышениями — “гороховидным”. Леггорны имеют простой или листовидный гребень, состоящий из одной вертикальной пластинки. Гибридологический анализ показал, что простой гребень ведет себя как полностью рецессивный признак по отношению к розовидному и гороховидному. Расщепление в F 2 соответствует формуле 3: 1. При скрещивании же между собой рас с розовидным и гороховидным гребнем у гибридов первого поколения развивается совершенно новая форма гребня, напоминающая половинку ядра грецкого ореха, в связи с чем гребень был назван “ореховидным”. При анализе второго поколения было установлено, что соотношение разных форм гребня в F 2 соответствует формуле 9: 3: 3: 1, что указывало на дигибридный характер скрещивания. Была разработана схема скрещивания, объясняющая механизм наследования этого признака.
В определении формы гребня у кур принимают участие два неаллельных гена. Доминантный ген R контролирует развитие розовидного гребня, а доминантный ген P — гороховидного. Комбинация рецессивных аллелей этих генов rrpp вызывает развитие простого гребня. Ореховидный гребень развивается при наличии в генотипе обоих доминантных генов.
Наследование формы гребня у кур можно отнести к комплементарному взаимодействию неаллельных генов. Комплементарными, или дополнительными, считаются гены, которые при совместном действии в генотипе в гомо- или гетерозиготном состоянии обусловливают развитие нового признака. Действие же каждого из генов в отдельности воспроизводит признак одного из родителей.
Схема, иллюстрирующая взаимодействие неаллельных генов,
определяющих форму гребня у кур
Наследование генов, определяющих форму гребня у кур, полностью укладывается в схему дигибридного скрещивания, так как они ведут себя при распределении независимо. Отличие от обычного дигибридного скрещивания проявляется только на уровне фенотипа и сводится к следующему:
- Гибриды F 1 не похожи ни на одного из родителей и обладают новым признаком;
- В F 2 появляются два новых фенотипических класса, которые являются результатом взаимодействия либо доминантных (ореховидный гребень), либо рецессивных (простой гребень) аллелей двух независимых генов.
Механизм комплементарного взаимодействия подробно изучен на примере наследования окраски глаз у дрозофилы. Красная окраска глаз у мух дикого типа определяется одновременным синтезом двух пигментов — бурого и ярко-красного, каждый из которых контролируется доминантным геном. Мутации, затрагивающие структуру этих генов, блокируют синтез либо того, либо другого пигмента. Так, рецессивная мутация brown (ген находится во 2-й хромосоме) блокирует синтез ярко-красного пигмента, в связи с чем у гомозигот по этой мутации бурые глаза. Рецессивная мутация scarlet (ген располагается в 3-й хромосоме) нарушает синтез бурого пигмента, и поэтому гомозиготы stst имеют ярко-красные глаза. При одновременном присутствии в генотипе обоих мутантных генов в гомозиготном состоянии не вырабатываются оба пигмента и глаза у мух белые.
В описанных примерах комплементарного взаимодействия неаллельных генов формула расщепления по фенотипу в F 2 соответствует 9: 3: 3: 1. Такое расщепление наблюдается в том случае, если взаимодействующие гены по отдельности имеют неодинаковое фенотипическое проявление и оно не совпадает с фенотипом гомозиготного рецессива. Если это условие не соблюдается, в F 2 имеют место иные соотношения фенотипов.
Например, при скрещивании двух разновидностей фигурной тыквы со сферической формой плода гибриды первого поколения обладают новым признаком — плоскими или дисковидными плодами. При скрещивании гибридов между собой в F 2 наблюдается расщепление в соотношении 9 дисковидных: 6 сферических: 1 удлиненная.

Анализ схемы показывает, что в определении формы плода принимают участие два неаллельных гена с одинаковым фенотипическим проявлением (сферическая форма). Взаимодействие доминантных аллелей этих генов дает дисковидную форму, взаимодействие рецессивных аллелей — удлиненную.
Еще один пример комплементарного взаимодействия дает наследование окраски шерсти у мышей. Дикая серая окраска определяется взаимодействием двух доминантных генов. Ген А отвечает за присутствие пигмента, а ген В — за его неравномерное распределение. Если в генотипе присутствует только ген А (А-bb ), то мыши равномерно окрашены в черный цвет. Если присутствует только ген В (ааВ- ), то пигмент не вырабатывается и мыши оказываются неокрашенными, так же как и гомозиготный рецессив ааbb . Такое действие генов приводит к тому, что в F 2 расщепление по фенотипу соответствует формуле 9: 3: 4.

F 2
| AB | Ab | aB | ab | |
| AB | AABB сер. |
AABb сер. |
AaBB сер. |
AaBb сер. |
| Ab | AABb сер. |
AAbb черн. |
AaBb сер. |
Aabb черн. |
| aB | AaBB сер. |
AaBb сер. |
aaBB бел. |
aaBb бел. |
| ab | AaBb сер. |
Aabb черн. |
aaBb бел. |
aabb |
Комплементарное взаимодействие описано также при наследовании окраски цветов у душистого горошка. Большая часть сортов этого растения имеет пурпурные цветы с фиолетовыми крыльями, которые характерны для дикой сицилийской расы, но есть также сорта с белой окраской. Скрещивая растения с пурпурной окраской цветов с растениями с белыми цветами Бетсон и Пеннет установили, что пурпурная окраска цветов полностью доминирует над белой, и в F 2 наблюдается соотношение 3: 1. Но в одном случае от скрещивания двух белых растений получилось потомство, состоящее только из растений с окрашенными цветами. При самоопылении растений F 1 было получено потомство, состоящее из двух фенотипических классов: с окрашенными и неокрашенными цветами в соотношении 9/16: 7/16.
Полученные результаты объясняются комплементарным взаимодействием двух пар неаллельных генов, доминантные аллели которых (С и Р ) в отдельности не способны обеспечить развитие пурпурной окраски, так же как и их рецессивные аллели (ссрр ). Окраска проявляется только при наличии в генотипе обоих доминантных генов, взаимодействие которых обеспечивает синтез пигмента.

пурп.
F 2
| CP | Cp | cP | cp | |
| CP | CCPP пурп. |
CCPp пурп. |
CcPP пурп. |
CcPp пурп. |
| Cp | CCPp пурп. |
CCpp бел. |
CcPp пурп. |
Ccpp бел. |
| cP | CcPP пурп. |
CcPp пурп. |
ccPP бел. |
ccPp бел. |
| cp | CcPp пурп. |
Ccpp бел. |
ccPp бел. |
В приведенном примере формула расщепления в F 2 — 9: 7 обусловлена отсутствием у доминантных аллелей обоих генов собственного фенотипического проявления. Однако такой же результат получается и в том случае, если взаимодействующие доминантные гены имеют одинаковое фенотипическое проявление. Например, при скрещивании двух сортов кукурузы с фиолетовой окраской зерновок в F 1 все гибриды имеют желтые зерновки, а в F 2 наблюдается расщепление 9/16 желт. : 7/16 фиол.
Эпистаз — другой тип неаллельного взаимодействия, при котором происходит подавление действия одного гена другим неаллельным ему геном. Ген, который препятствует проявлению другого гена, называется эпистатичным, или супрессором, а тот, чье действие подавляется, гипостатичным. В качестве эпистатичного гена может выступать как доминантный, так и рецессивный ген (соответственно доминантный и рецессивный эпистаз).
Примером доминантного эпистаза служит наследование окраски шерсти у лошадей и окраски плодов у тыквы. Схема наследования этих двух признаков абсолютно одинаковая.

F 2
| CB | Cb | cB | cb | |
| CB | CCBB сер. |
CCBB сер. |
CcBB сер. |
CcBb сер. |
| Cb | CCBb сер. |
CCbb сер. |
CcBb сер. |
Ccbb сер. |
| cB | CcBB сер. |
CcBb сер. |
ccBB черн. |
ccBb черн. |
| cb | CcBb сер. |
Ccbb сер. |
ccBb черн. |
ccbb рыж. |
Из схемы видно, что доминантный ген серой окраски С является эпистатичным по отношению к доминантному гену В , который обусловливает черную окраску. В присутствии гена С ген В своего действия не проявляет, и поэтому гибриды F 1 несут признак, определяемый эпистатичным геном. В F 2 класс с обоими доминантными генами сливается по фенотипу (серая окраска) с классом, у которого представлен только эпистатичный ген (12/16). Черная окраска проявляется у 3/16 гибридных потомков, в генотипе которых отсутствует эпистатичный ген. В случае гомозиготного рецессива отсутствие гена-супрессора позволяет проявиться рецессивному гену с, который вызывает развитие рыжей окраски.

Доминантный эпистаз описан также при наследовании окраски пера у кур. Белый цвет оперенья у кур породы леггорнов доминирует над окрашенным черных, рябых и других цветных пород. Однако белая окраска других пород (например, плимутроков) рецессивна по отношению к цветному оперению. Скрещивания между особями с доминантной белой окраской и особями с рецессивной белой окраской в F 1 дают белое потомство. В F 2 наблюдается расщепление в соотношении 13: 3.
Анализ схемы показывает, что в определении окраски пера у кур принимают участие две пары неаллельных генов. Доминантный ген одной пары (I ) является эпистатичным по отношению к доминантному гену другой пары, вызывающему развитие окраски (C ). В связи с этим окрашенное оперение имеют только те особи, в генотипе которых присутствует ген С , но отсутствует эпистатичный ген I . У рецессивных гомозигот ссii отсутствует эпистатичный ген, но у них нет гена, который обеспечивает выработку пигмента (C ), поэтому они имеют белую окраску.
В качестве примера рецессивного эпистаза можно рассмотреть ситуацию с геном альбинизма у животных (см. выше схему наследования окраски шерсти у мышей). Присутствие в генотипе двух аллелей гена альбинизма (аа ) не дает возможности проявиться доминантному гену окраски (B ) — генотипы ааВ- .
Полимерный тип взаимодействия был впервые установлен Г. Нильсеном-Эле при изучении наследования окраски зерна у пшеницы. При скрещивании краснозерного сорта пшеницы с белозерным в первом поколении гибриды были окрашенными, но окраска была розовой. Во втором поколении только 1/16 часть потомства имела красную окраску зерна и 1/16 — белую, у остальных окраска была промежуточной с разной степенью выраженности признака (от бледно-розовой до темно-розовой). Анализ расщепления в F 2 показал, что в определении окраски зерна участвуют две пары неаллельных генов, действие которых суммируется. Степень выраженности красной окраски зависит от количества доминантных генов в генотипе.
Полимерные гены принято обозначать одинаковыми буквами с добавлением индексов, в соответствии с числом неаллельных генов.
Действие доминантных генов в данном скрещивании является аддитивным, так как добавление любого из них усиливает развитие признака.

F 2
| A 1 A 2 | A 1 a 2 | a 1 A 2 | a 1 a 2 | |
| A 1 A 2 | A 1 A 1 A 2 A 2 красн. |
A 1 A 1 A 2 Aa 2 ярко-розов. |
A 1 a 1 A 2 A 2 ярко-розов. |
A 1 a 1 A 2 a 2 розов. |
| A 1 a 2 | A 1 A 1 A 2 a 2 ярко-розов. |
A 1 A 1 a 2 a 2 розов. |
A 1 a 1 A 2 a 2 розов. |
A 1 a 1 a 2 a 2 бледно-розов. |
| a 1 A 2 | A 1 a 1 A 2 A 2 ярко-розов. |
A 1 a 1 A 2 a 2 розов. |
a 1 a 1 A 2 A 2 розов. |
a 1 a 1 A 2 a 2 бледно-розов. |
| a 1 a 2 | A 1 a 1 A 2 a 2 розов. |
A 1 a 1 a 2 a 2 бледно-розов. |
a 1 a 1 A 2 a 2 бледно-розов. |
a 1 a 1 a 2 a 2 |
Описанный тип полимерии, при котором степень развития признака зависит от дозы доминантного гена, называется кумулятивным. Такой характер наследования обычен для количественных признаков, к которым следует отнести и окраску, т.к. ее интенсивность обусловлена количеством вырабатываемого пигмента. Если не учитывать степень выраженности окраски, то соотношение окрашенных и неокрашенных растений в F 2 соответствует формуле 15: 1.
Однако в некоторых случаях полимерия не сопровождается кумулятивным эффектом. В качестве примера можно привести наследование формы семян у пастушьей сумки. Скрещивание двух рас, одна из которых имеет треугольные плоды, а другая яйцевидные дает в первом поколении гибриды с треугольной формой плода, а во втором поколении наблюдается расщепление по этим двум признакам в соотношении 15 треуг. : 1 яйцев.
Данный случай наследования отличается от предыдущего только на фенотипическом уровне: отсутствие кумулятивного эффекта при увеличении дозы доминантных генов обусловливает одинаковую выраженность признака (треугольная форма плода) независимо от их количества в генотипе.
К взаимодействию неаллельных генов относят также явление плейотропии — множественного действия гена, влияния его на развитие нескольких признаков. Плейотропное действие генов является результатом серьезного нарушения обмена веществ, обусловленного мутантной структурой данного гена.
Так, например, ирландские коровы породы декстер отличаются от близкой по происхождению породы керри укороченными ногами и головой, но одновременно лучшими мясными качествами и способностью к откорму. При скрещивании коров и быков породы декстер 25% телят имеют признаки породы керри, 50% сходны с породой декстер, а в остальных 25% случаев наблюдаются выкидыши уродливых бульдогообразных телят. Генетический анализ позволил установить, что причиной гибели части потомства является переход в гомозиготное состояние доминантной мутации, вызывающей недоразвитие гипофиза. В гетерозиготе этот ген приводит к появлению доминантных признаков коротконогости, короткоголовости и повышенной способности к отложению жира. В гомозиготе этот ген имеет летальный эффект, т.е. в отношении гибели потомства он ведет себя как рецессивный ген.
Летальный эффект при переходе в гомозиготное состояние характерен для многих плейотропных мутаций. Так, у лисиц доминантные гены, контролирующие платиновую и беломордую окраски меха, не оказывающие летального действия в гетерозиготе, вызывают гибель гомозиготных зародышей на ранней стадии развития. Аналогичная ситуация имеет место при наследовании серой окраски шерсти у овец породы ширази и недоразвития чешуи у зеркального карпа. Летальный эффект мутаций приводит к тому, что животные этих пород могут быть только гетерозиготными и при внутрипородных скрещиваниях дают расщепление в соотношении 2 мутанта: 1 норма.

F 1
F 1: 2 плат. : 1 черн.
Однако большинство летальных генов рецессивны, и гетерозиготные по ним особи имеют нормальный фенотип. О наличии у родителей таких генов можно судить по появлению в потомстве гомозиготных по ним уродов, абортусов и мертворожденных. Чаще всего подобное наблюдается в близкородственных скрещиваниях, где родители обладают сходными генотипами, и шансы перехода вредных мутаций в гомозиготное состояние достаточно высоки.
Плейотропные гены с летальным эффектом есть у дрозофилы. Так, доминантные гены Curly — загнутые вверх крылья, Star — звездчатые глаза, Notch — зазубренный край крыла и ряд других в гомозиготном состоянии вызывают гибель мух на ранних стадиях развития.
Известная рецессивная мутация white , впервые обнаруженная и изученная Т. Морганом, также имеет плейотропный эффект. В гомозиготном состоянии этот ген блокирует синтез глазных пигментов (белые глаза), снижает жизнеспособность и плодовитость мух и видоизменяет форму семенников у самцов.
У человека примером плейотропии служит болезнь Марфана (синдром паучьих пальцев, или арахнодактилия), которая вызывается доминантным геном, вызывающим усиленный рост пальцев. Одновременно он определяет аномалии хрусталика глаза и порок сердца. Болезнь протекает на фоне повышения интеллекта, в связи с чем ее называют болезнью великих людей. Ею страдали А. Линкольн, Н. Паганини.
Плейотропный эффект гена, по всей видимости, лежит в основе коррелятивной изменчивости, при которой изменение одного признака влечет за собой изменение других.
К взаимодействию неаллельных генов следует отнести также влияние генов-модификаторов, которые ослабляют или усиливают функцию основного структурного гена, контролирующего развитие признака. У дрозофилы известны гены-модификаторы, модифицирующие процесс жилкования крыльев. Известно не менее трех генов-модификаторов, влияющих на количество красного пигмента в волосе крупного рогатого скота, в результате чего окраска шерсти у разных пород колеблется от вишневой до палевой. У человека гены-модификаторы изменяют окраску глаз, усиливая или ослабляя ее интенсивность. Их действием объясняется разная окраска глаз у одного человека.
Существование явления взаимодействия генов привело к появлению таких понятий, как “генотипическая среда” и “генный баланс”. Под генотипической средой подразумевается то окружение, в которое попадает вновь возникающая мутация, т.е. весь комплекс генов, имеющихся в данном генотипе. Понятие “генный баланс” касается соотношения и взаимодействия между собой генов, влияющих на развитие признака. Обычно гены обозначают названием признака, возникающего при мутации. На самом же деле проявление этого признака часто является результатом нарушения функции гена под влиянием других генов (супрессоров, модификаторов и др.). Чем сложнее генетический контроль признака, чем больше генов участвуют в его развитии, тем выше наследственная изменчивость, так как мутация любого гена нарушает генный баланс и приводит к изменению признака. Следовательно, для нормального развития особи необходимо не только присутствие генов в генотипе, но и осуществление всего комплекса межаллельных и неаллельных взаимодействий.
Изучение закономерностей наследования признаков, сцепленных с полом, и выяснение возможностей их использования в практической деятельности человека можно проводить только на животных с ярко выраженным половым диморфизмом, имеющих характерные признаки, гены которых находятся в половой Х-хромосоме. В условиях пришкольного участка для этих целей наиболее удобно использовать кур, канареек и других животных.
План опыта с курами
Тема опыта. Закономерности наследования окраски оперения у кур.
Задачи опыта. 1. Закрепить навыки ухода за курами. 2. Установить специфику наследования признаков, гены которых находятся в половых хромосомах. 3. Выяснить возможности использования в практической деятельности человека закономерностей наследования признаков, гены которых находятся в половых хромосомах.
Выбор и содержание исходных пар. Для опыта в качестве родительских форм отобрать молодых здоровых гомозиготных птиц двух пород: с полосатым оперением (плимутрок) и с черным оперением (австралорпы, украинские черные или др.). Опыт провести в двух вариантах: 1) прямое скрещивание (куры черные´петух полосатый); 2) обратное скрещивание (куры полосатые´петух черный). В каждом варианте взять по 2-3 курицы и одному петуху. Данные о родителях записать в журнал гибридизации по следующей схеме:
| Дата | Родители, гибриды | Пол | Прямое скрещивание | Обратное скрещивание | ||
| количество особей | окраска оперения | количество особей | окраска оперения | |||
| Р | Куры | |||||
| Петухи | ||||||
| F 1 | Куры | |||||
| Петухи | ||||||
| F 2 | Куры | |||||
| Петухи |
Для получения отобранных для опыта птиц поместить в клетки отдельно по вариантам и содержать при одинаковом уходе и кормлении (обычных для кур). Снесенные яйца учитывать, хранить и помещать для насиживания отдельно по вариантам, снабдив этикеткой, в которой указана гибридная комбинация.
Гибриды F 1 . Выращивать гибридных цыплят нужно отдельно по вариантам в обычных для кур условиях кормления и содержания. Когда цыплята достигнут половой зрелости, в каждом варианте установить окраску оперения у курочек и петушков, подсчитать количество особей с одинаковым фенотипом, данные наблюдений занести в журнал гибридизации. Для получения гибридов F 2 в каждом варианте из гибридов F 1 отобрать по 2-3 курочки и одному петушку, поместить в клетки и содержать так же, как родителей. Яйца хранить и помещать для насиживания отдельно по вариантам с этикеткой, в которой указана комбинация.
Гибриды F 2 . Выращивать цыплят из F 2 нужно отдельно по вариантам, как гибриды F 1 . Когда они достигнут половой зрелости, в каждом варианте подсчитать количество курочек и петушков, определить окраску оперения. Данные наблюдений внести в журнал гибридизации.
Анализ результатов опыта. Данные учетов проанализировать и сделать выводы о характере окраски оперения у кур.
При правильном ведении опыта в разных вариантах характер наследования окраски оперения у птиц должен быть иным. При прямом скрещивании (1-й вариант) в F 1 все курочки и петушки должны быть полосатыми. Это говорит о том, что ген полосатости доминирует над геном черной окраски. В F 2 все петушки должны быть полосатыми, а из курочек 50% полосатыми и 50% черными. При обратном скрещивании в F 1 все петушки будут черными, как мать, а курочки - полосатыми, как отец; в F 2 50% петушков и 50% курочек будут черными, а 50% -полосатыми.
Такой характер наследования говорит о том, что у кур ген, определяющий окраску оперения, находится в половой хромосоме. (У птиц гетерогаметный пол женский. Половой комплекс самки - XX , самца - XY ). Схема наследования этого признака приведена на рисунке 75.
Данные опыта использовать на уроках общей биологии в X классе при изучении сцепленного с полом наследования и в практике птицеводства, для определения пола молодых цыплят, которые, как известно, в раннем возрасте не имеют внешне заметных половых различий, а вместе с тем экономически целесообразно сразу после рождения отделить петушков и курочек и определить им различные режимы кормления и содержания, так как в дальнейшем курочки пополнят стадо несушек, а петушки будут использованы как бройлеры.
Опыт с канарейками. Сцепленное с полом наследование можно продемонстрировать и на других объектах, например на канарейках, у которых доминантный ген А , определяющий зеленое оперение, и его рецессивная аллель (а ), определяющая коричневое оперение, находятся в половой (X ) хромосоме. Скрещивание, как и у кур, провести в двух вариантах: 1) прямое скрещивание (зеленый (АА )´коричневая (ау ); 2) обратное скрещивание (коричневый (аа )´зеленая (Ау ).
4 . 2 . ЭПИСТАЗ
Эпистаз (от греч. epístasis – остановка, препятствие), взаимодействие двух неаллельных (т. е. относящихся к разным локусам) генов, при котором один ген, называемый эпистатичным или геномсупрессором, подавляет действие другого гена, называемого гипостатичным. Гены-супрессоры известны у животных (млекопитающих, птиц, насекомых) и у растений. Обычно они обозначаютсяI илиSu в случае доминантного состояния генов иi илиsu для их рецессивных аллелей (от английских словinhibitor илиsupressor ). При эпистазе аллель одного из генов подавляет действие аллелей других генов, напримерА >В илиВ >А ,а >В илиb >А и т. д.
Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному. В случае комплементарного взаимодействия происходит дополнение одного гена другим. Эпистатическое действие генов по своему характеру очень похоже на явление доминирования, разница состоит лишь в том, что при доминировании аллель подавляет проявление рецессивного аллеля, принадлежащего той же самой аллеломорфной паре. При эпистазе же аллель одного гена подавляет проявление аллеля из другой аллеломорфной пары, т. е. неаллельного гена.
Фенотипически эпистаз выражается в отклонении от расщепления, ожидаемого при дигенном наследовании, однако нарушения законов Г. Менделя в этом случае нет, так как распределение аллелей взаимодействующих генов полностью соответствует закону независимого комбинирования признаков.
В настоящее время эпистаз разделяют на два типа: доминантный и рецессивный. Наиболее известные примеры взаимодействия генов по типу эпистаза приведены в табл. 13.
Таблица 13
Расщепление признаков при эпистазе |
||
Расщепление в F 2 | ||
Наследование окраски оперения у кур. |
||
Наследование окраски плодов у тыквы (белые, желтые и |
||
зеленые). |
||
Наследование окраски зерен овса (черные, серые и белые). |
||
Наследование масти лошадей (серых, вороных и рыжих). |
||
Наследование окраски шерсти у собак породы Лабрадор. |
||
Наследование окраски шерсти у мышей. |
||
Двойной рецессивный эпистаз (криптомерия). |
||
Доминантный эпистаз (А >В илиВ >А ). Под доминантным эпистазом понимают подавление доминантным аллелем одного гена действия аллельной пары другого гена.
Эпистатическая система обнаружена у кур. Некоторые породы кур имеют белое оперение (белый леггорн, белый плимутрок, виандотт и др.), другие же породы имеют окрашенное оперение (австралорп, ньюгемпшир, полосатый плимутрок и др.). Белое оперение разных пород кур определяется несколькими различными генами. Так, например, доминантная белая окраска определяется генами CCII (белые леггорны), а рецессивная белая –ccii (белые виандотты). ГенС определяет наличие предшественника пигмента (хромогена), т. е. окрашенность пера, его аллельс – отсутствие хромогена и, следовательно, неокрашенность пера птицы. ГенI является подавителем действия генаС , аллельi не подавляет его действия. В присутствии даже одной дозы генаI в генотипе птицы действие генов окраски не проявится. Поэтому при скрещивании белых леггорнов (CCII ) с белыми виндоттами (ccii) гибридыF 1 оказываются белыми (СсIi ). При скрещивании между собой гибридовF 1 во втором поколении имеет место расщепление по окраске в отношении 13 белых: 3 окрашенных (рис. 38).
F1 : | ||
F2 : | 9 С -I - : 3 ссI - : | l ccii: 3 С-ii |
окрашенные |
Рис. 38 . Наследование окраски у кур при взаимодействии двух пар генов (эпистаз):I – подавляет окраску,i –не подавляет окраску,
C – наличие пигмента,c –отсутствие пигмента

Таким образом, подавление действия доминантного аллеля гена, определяющего развитие окраски (С ), доминантным аллелем другого гена (I ) обусловливает вF 2 расщепление по фенотипу 13: 3.
Доминантный эпистаз может давать и другое соотношение фенотипов в F 2 , а именно: 12: 3: 1. В этом случае рецессивная гомозигота (aabb) фенотипически отличима от одного из гетерозиготных классов А-bb или ааВ- . Такой тип расщепления признаков установлен для окраски плодов у тыквы (рис. 39). У этого растения известно три цвета плодов: белая, желтая и зеленая. Доминантный аллель генаА определяет желтую окраску тыквы, рецессивный аллель – зеленый цвет. Второй генВ проявляет эпистатическое действие – подавляет образование пигмента как желтого, так и зеленого цвета, делая их белыми. Рецессивный аллельв – не влияет на проявление окраски плодов тыквы. При скрещивании растений с белыми (ААВВ ) и зелеными (ааbb ) плодами, все потомствоF 1 будет белым. ВF 2 расщепление признаков будет соответствовать формуле 12: 3: 1.
F1 : А-В-
Рис. 39 . Наследование признаков окраски плодов у тыквы
В приведенном примере ген-подавитель не определяет сам какойлибо качественной реакции или синтетического процесса, а лишь подавляют действие других генов.
Несколько иной механизм доминантного эпистаза известен для окраски зерна у овса. В этом случае ген-подавитель выполняет две функции – обеспечивает проявление признака и одновременно про-
являет эпистатическое действие в отношении другого гена. У этой культуры были установлены доминантные гены, определяющие черную (ген А ) и серую (генВ ) окраску зерна. Кроме того, генА проявляет эпистатическое действие в отношении генаВ . При скрещивании родительских форм черносемянных (ААВВ ) и белосемянных (ааbb ) вF 1 все потомство будет черносемянным (АаВb ). Так как генА подавляет проявление генаВ, в F 1 все потомство будет черносемянное. ВF 2 расщепление будет 12: 3: 1:
F 2: | 9 А-В- | : 3 А-bb : | 3 ааВ- | : 1 ааbb |
|
12 черные: | |||||
Еще одним примером доминантного эпистаза является взаимодействие генов, определяющих окраску шерсти у лошадей. Ген В в доминантном состоянии определяет вороную масть, а в рецессивном
– рыжую. Ген С в доминантном состоянии определяет серую масть (вызывает раннее поседение лошадей). Кроме того, дополнительно ген С проявляет эпистатическое действие в отношении генаВ независимо от того, находится ли последний в доминантном или рецессивном состоянии. В результате действия гена подавителяС масть лошадей независимо от аллельного состояния генаВ становиться серой. Поэтому от скрещивания серых лошадей генотипаВВСС с рыжими (bbсс ) вF 1 рождается серое потомство (ВbСс ). При скрещивании лошадей серой масти между собой вF 2 наблюдается расщеп-
ление 12: 3: 1.
F1 : | ||||
F2 : | 9 В-С- : | 3 bbС- | : 3 В-сс : | 1 ааbb |
12 серые: 3 вороные: 1 рыжие
Рецессивный эпистаз. Под рецессивным эпистазом понимают такой тип взаимодействия, когда рецессивный аллель одного гена, будучи в гомозиготном состоянии, не дает возможности проявиться
доминантному или рецессивному аллелям другого гена: аа >В;
аа> bbили bb> А; bb> аа.
Мы уже имели возможность познакомиться с расщеплением 9:3:4 как результатом комплементарного взаимодействия генов. Но эти же случаи можно рассматривать и как пример рецессивного эпистаза.
Примером рецессивного эпистаза является окраска шерсти у собак породы лабрадор. Пигментация шерсти обеспечивается геном В, который в доминантном состоянии дает черную масть, а в рецессивном (b ) – коричневую. Имеется также генЕ , который в доминантном состоянии не влияет на проявление окраски, но будучи в рецессивном состоянии (сс ) подавляет синтез пигмента как черного, так и коричневого. Такие собаки становятся белыми. Расщепление в
F 2 будет следующее: | ||||
F2 : 9 В-Е- | ввЕ- : | 3 В-ее | : 1 ввее |
|
коричневые | ||||
9 черные: 3 коричневые: 4 белые
При скрещивании черных мышей (AAсс ) с альбиносами (ааСС ) все особиF 1 (AaСс ) имеют окраску типа агути (пример комплементарного действия генов), а вF 2 9 частей всех особей оказываются агути (А -С -), 3 части черные (А -сс ) и 4 – альбиносы (ааС - иaaсс ). Эти результаты можно объяснить, предположив, что имеет место рецессивный эпистаз типааа >С -. При этом мыши генотипаааС - оказываются белыми, потому что гена в гомозиготном состоянии, обусловливая отсутствие пигмента, препятствует тем самым проявлению гена-распределителя пигментаС .
Интересным примером рецессивного эпистаза является фенотип крови Бомбей. В редких случаях у жителей Бомбея (Индия), в генотипе которых имеются доминантные аллели I А илиI B , группа крови оказывается нулевой (О). Было установлено, что этот дефект обусловлен рецессивной мутацией в генеН , не гомологичном локусамА илиВ, которая в гомозиготном состоянии приводит к тому, что в процессе синтезаА - или В -антигенов образуется дефектное Н -вещество, с которым фермент гликозилтрансфераза взаимодействовать не может и, следовательно, нормальные антигены не образуются. Природа вещества Н -вещества в настоящее время еще не исследова-
на. Антигены А иВ обнаруживаются только у лиц с генотипомНН илиHh . Когда оба родителя имеют генотипI А I B Нh, среди потомков могут встретиться различные группы крови в следующем соотноше-
нии: 3А: 6АВ: 3В: 4О.
Кроме описанных случаев рецессивного эпистаза, существуют и такие, когда рецессивный аллель каждого из генов в гомозиготном состоянии одновременно реципрокно подавляет действие другой пары генов, т.е. аа эпистатирует над В -, abb над А -. Такое взаимодействие двух рецессивных генов называют двойным рецессивным эпистазом (криптомерией). При этом в дигибридном скрещивании расщепление по фенотипу будет соответствовать 9: 7, как и в случае комплементарного взаимодействия генов. Следовательно, одно и то же отношение можно трактовать и как комплементарное взаимодействие, и как эпистатирование. Сам по себе генетический анализ наследования при взаимодействии генов без учета биохимии и физиологии развития признака в онтогенезе не может раскрыть природы этого взаимодействия. Но без генетического анализа нельзя понять наследственной детерминации развития этих признаков.
4 . 3 . ПОЛИМЕРИЯ
В рассмотренных до сих пор типах взаимодействия генов мы касались альтернативных, т. е. качественно различающихся, признаков. Однако такие свойства организмов, как рост, масса, яйценоскость кур, количество молока и его жирность у скота, длина шерсти у овец, количество белка в эндосперме зерна кукурузы и пшеницы, содержание витаминов в растениях, скорость протекания биохимических реакций, свойства нервной деятельности животных и т. п., нельзя разложить на четкие фенотипические классы. Такие признаки необходимо оценивать в количественном выражении, поэтому они чаще всего называются количественными, или мерными.
Изучение наследования полимерных признаков было начато в первом десятилетии нашего столетия. Так, при скрещивании растений пшеницы с красными и белыми (неокрашенными) зернами шведский генетик Г. Нильсон-Эле в 1908 г. обнаружил вF 2 обычное моногибридное расщепление в отношении 3: 1. Однако при скрещивании некоторых линий пшеницы, различающихся по таким же признакам, вF 2 наблюдается расщепление в отношении 15/16 окрашенных и 1/16 белых (рис. 40).
Окраска зерен из первой группы варьировала от темно-красных до бледно-красных. Генетический анализ растений пшеницы вF 3 из семян

F 2 , что растения, выращенные из белых зерен и из зерен с наиболее темной (красной) окраской, в дальнейшем не дают расщепления. Из зерен с окраской промежуточного типа развились растения, давшие в последующих поколениях расщепление по окраске зерна.
A1 A1 A2 A2 | a1 a1 a2 a2 |
|||
F3 :
A 1A 2♂
A1 а2
а1 A2
а1 а2
A1 а1 A2 а2
♀ A 1 A 2 | A1 а2 | а1 A2 | а1 а2 |
A1 A1 | A1 A1 | A1 а1 | A1 а1 |
А2 А2 | А2 а2 | А2 А2 | А2 а2 |
A1 A1 | A1 A1 | A1 а1 | A1 а1 |
А2 а2 | а2 а2 | А2 а2 | а2 а2 |
A1 а1 | A1 а1 | а1 а1 | а1 а1 |
А2 А2 | А2 а2 | А2 А2 | А2 а2 |
A1 а1 | A1 а1 | а1 а1 | а1 а1 |
А2 а2 | а2 а2 | А2 а2 | а2 а2 |
Рис. 40 . Наследование окраски зерна уTriticum при взаимодействии двух пар генов (полимерия).
Анализ характера расщепления позволил установить, что в данном случае красную окраску зерен определяют два доминантных аллеля двух разных генов, а сочетание их рецессивных аллелей в гомозиготном состоянии определяет отсутствие окраски. Интенсивность окраски зерен зависит от числа доминантных генов, присутствующих в генотипе.

Полимерия присуща генам, которые представлены самостоятельными единицами, т.е. являются неаллельными, однако их продукты выполняют одну и ту же функцию (рис. 41).
Ген А 1
Такие гены были названы полимерными, и, поскольку однозначно влияют на один и тот же признак было принято обозначать их одной латинской буквой с указанием индекса для разных членов: А 1 ,А 2 ,А 3 и т.д. Следовательно, исходные родительские формы, давшие расщепление вF 2 15: 1, имели генотипыА 1 А 1 А 2 А 2 иа 1 а 1 а 2 а 2 .
Очевидно, что при тригибридном скрещивании, если у гибрида F 1 число полимерных генов в гетерозиготном состоянии оказывается не
два, а три А 1 а 1 А 2 а 2 А 3 а 3 или более, то число |
||||||||||
комбинаций генотипов в F 2 увеличивается. |
||||||||||
В опыте Г. Нильсона-Эле тригибридное |
||||||||||
расщепление в F 2 по генам окраски зерен |
||||||||||
пшеницы давало соотношение: 63 растения |
||||||||||
с красным эндоспермом и 1 растение – с не- |
||||||||||
окрашенным. В F 2 наблюдались все перехо- |
||||||||||
ды от интенсивной окраски зерен с геноти- |
||||||||||
пом А 1 А 1 А 2 А 2 А 3 А 3 до полного ее отсутствия |
||||||||||
у а 1 а 1 а 2 а 2 а 3 а 3 . При этом частоты генотипов с |
||||||||||
разным количеством | доминантных | |||||||||
распределились | следующий | |||||||||
1+6+15+20+15+6+1=64. | ||||||||||
Рис. 42 . Кривые распределения | На рис. 42 приведены кривые рас- |
|||||||||
частот генотипов в F2 | пределения частот | генотипов с разным |
||||||||
кумулятивной полимерии при | числом доминантных генов кумулятивно- |
|||||||||
скрещивании: | ||||||||||
1 – моногибридном; 2 – диги- | го действия при независимом их сочета- |
|||||||||
бридном; 3 – тригибридном | нии в моно-, ди- и тригибридном скрещи- |
|||||||||
ваниях. Из этого сопоставления видно, что чем большее число доминантных генов определяет данный признак, тем больше амплитуда изменчивости.
При наследовании количественного признака потомство гибрида образует непрерывный вариационный ряд по фенотипическому проявлению данного признака.
Изучение полимерных генов имеет не только теоретический, но и большой практический интерес. Установлено, что многие хозяйственно ценные признаки у животных и растений, такие как молочность скота, яйценоскость кур, вес и рост животного, длина колоса, длина початка кукурузы, содержание сахара в корнеплодах свеклы, плодовитость и скороспелость животных, длина вегетационного периода у растений и многие другие наследуются по типу полимерии.
Изменчивость количественного признака в отличие от альтернативного оценивается амплитудой его варьирования. Сама амплитуда варьирования признака наследственно определена и имеет приспособительное значение в индивидуальном развитии. В качестве примера к сказанному приведем опыт Е. Иста по скрещиванию двух форм кукурузы – длиннопочатковой и короткопочатковой. Как видно из результатов, представленных на рис. 43, початки по их длине у исходных линий кукурузы № 60 (короткопочатковая) и № 54 (длиннопочатковая), а также у гибридов первого и второго поколений распределяются с определенной закономерностью. Нетрудно заметить, что эти две линии сильно различаются между собой, но в пределах каждой из них длина початков колеблется незначительно. Это указывает на то, что они наследственно сравнительно однородны. Захождения в размерах початков у родительских форм нет.
У гибридных растений длина початков оказывается промежуточной, с небольшой изменчивостью ряда. При самоопылении растений F 1 в следующем поколении (F 2 ) размах изменчивости по длине початков значительно увеличивается. Если вычертить кривую распределения классов по длине початков, отложив на абсциссе размер початков, а на ординате – их количество, она оказывается сходной с кривой распределения полимерных доминантных генов (см. рис.42). Следовательно, непрерывный ряд изменений по длине початка кукурузы можно представить как ряд генотипов при тригибридном скрещивании с различным числом доминантных генов, обусловливающих данный количественный признак.

F1 :
F2 :
Рис. 43. Наследование и изменчивость длины початков (в сантиметрах)
у Zea mays в F 1 иF 2
Тот факт, что при небольшом числе исследованных растений второго поколения у некоторых из них воспроизводится длина початков, свойственная родительским формам, может указывать на участие небольшого числа полимерных генов в определении длины початка у скрещиваемых форм. Такое предположение вытекает из известной нам формулы 4n , определяющей число возможных комбинаций гамет, образующих зиготы вF 2 , в зависимости от числа пар генов, по которым различались исходные родительские формы. Появление вF 2 растений, сходных с родительскими формами, при объеме выборки, равном 221 растению, указывает на то, что число независимо наследующихся генов, определяющих длину початка, не должно превышать трех (43 = 64) или четырех (44 = 256).
Приведенные примеры анализа наследования количественных признаков иллюстрируют лишь один из возможных путей изучения сложных и колеблющихся в своем проявлении признаков. Большая изменчивость признака, прежде всего, указывает на его сложную генетическую обусловленность и, напротив, меньшая изменчивость признака – на меньшее число факторов его определяющих.