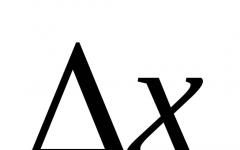Наружное коленчатое тело. Строение наружного коленчатого тела
Забугорье или метаталамус
Метаталамус (лат. Metathalamus) - часть таламической области головного мозга млекопитающих. Образован парными медиальным и латеральным коленчатыми телами, лежащими позади каждого таламуса.
Медиальное коленчатое тело находится позади подушки таламуса, оно, наряду с нижними холмиками пластинки крыши среднего мозга (четверохолмия), является подкорковым центром слухового анализатора. Латеральное коленчатое тело расположено книзу от подушки. Вместе с верхними холмиками пластинки крыши оно является подкорковым центром зрительного анализатора. Ядра коленчатых тел связаны проводящими путями с корковыми центрами зрительного и слухового анализаторов.
В медиальной части таламуса различают медиодорсалъное ядро и группу ядер средней линии.
Медиодорсальное ядро имеет двусторонние связи с обонятельной корой лобной доли и поясной извилиной больших полушарий, миндалевидным телом и переднемедиальным ядром таламуса. Функционально оно тесно связано также с лимбической системой и имеет двусторонние связи с корой теменной, височной и островковой долей мозга.
Медиодорсальное ядро участвует в реализации высших психических процессов. Его разрушение приводит к снижению беспокойства, тревожности, напряженности, агрессивности, устранению навязчивых мыслей.
Ядра средней линии многочисленны и занимают наиболее медиальное положение в таламусе. Они получают афферентные (т.е. восходящие) волокна от гипоталамуса, от ядер шва, голубого пятна ретикулярной формации ствола мозга и частично от спинно-таламических путей в составе медиальной петли. Эфферентные волокна от ядер средней линии направляются к гиппокампу, миндалевидному телу и поясной извилине больших полушарий, входящих в состав лимбической системы. Связи с корой больших полушарий двусторонние.
Ядра средней линии играют важную роль в процессах пробуждения и активации коры больших полушарий, а также в обеспечении процессов памяти.
В латеральной (т.е. боковой) части таламуса располагаются дорсолатералъная, вентролатеральная, вентральная заднемедиальная и задняя группы ядер.
Ядра дорсолатералъной группы относительно мало изучены. Известно, что они причастны к системе восприятия боли.
Ядра вентролатералъной группы анатомически и функционально различаются между собой. Задние ядра вентролатеральной группы часто рассматриваются как одно вентролатеральное ядро таламуса. Эта группа получает волокна восходящего пути общей чувствительности в составе медиальной петли. Сюда приходят также волокна вкусовой чувствительности и волокна от вестибулярных ядер. Эфферентные волокна, начинающиеся от ядер вентролатеральной группы, направляются в кору теменной доли больших полушарий, куда проводят соматосенсорную информацию от всего тела.
К ядрам задней группы (ядра подушки таламуса) идут афферентные волокна от верхних холмиков четверохолмия и волокна в составе зрительных трактов. Эфферентные волокна широко распространяются в коре лобной, теменной, затылочной, височной и лимбической долей больших полушарий.
Ядерные центры подушки таламуса причастны к комплексному анализу различных сенсорных раздражителей. Они играют значительную роль в перцептивной (связанной с восприятием) и когнитивной (познавательной, мыслительной) деятельности мозга, а также в процессах памяти - хранения и воспроизведения информации.
Интраламинарная группа ядер таламуса лежит в толще вертикальной Y-образной прослойки белого вещества. Интраламинарные ядра взаимосвязаны с базальными ядрами, зубчатым ядром мозжечка и корой больших полушарий.
Эти ядра играют важную роль в активационной системе мозга. Повреждение интраламинарных ядер в обоих таламусах приводит к резкому снижению двигательной активности, а также апатии и разрушению мотивационной структуры личности.
Кора больших полушарий благодаря двусторонним связям с ядрами таламуса способна оказывать регулирующее воздействие на их функциональную активность.
Таким образом, основными функциями таламуса являются:
переработка сенсорной информации от рецепторов и подкорковых переключающих центров с последующей передачей её коре;
участие в регуляции движений;
обеспечение связи и интеграции различных отделов мозга
Коленчатое тело
(corpus geniculatum)
Коле́нчатое те́ло латера́льное
(с. g. laterale, BNA, JNA) - К. т., лежащее на нижней поверхности таламуса латерально ручки верхнего холмика четверохолмия; место расположения подкоркового центра зрения. Коле́нчатое те́ло медиа́льное
(с. g. mediale, PNA, BNA, JNA) - К. т., расположенное кпереди и латеральнее ручки нижнего холмика четверохолмия; место расположения подкоркового центра слуха.
1. Малая медицинская энциклопедия. - М.: Медицинская энциклопедия. 1991-96 гг. 2. Первая медицинская помощь. - М.: Большая Российская Энциклопедия. 1994 г. 3. Энциклопедический словарь медицинских терминов. - М.: Советская энциклопедия. - 1982-1984 гг .
Смотреть что такое "Коленчатое тело" в других словарях:
- (corpus geniculatlim) общее название валикоподобных образований промежуточного мозга, составляющих метаталамус … Большой медицинский словарь
- (с. g. laterale, PNA, BNA, JNA) К. т., лежащее на нижней поверхности таламуса латерально от ручки верхнего холмика четверохолмия: место расположения подкоркового центра зрения … Большой медицинский словарь
- (с. g. mediale, PNA, BNA, JNA) К. т., расположенное кпереди и латеральнее ручки нижнего холмика четверохолмия; место расположения подкоркового центра слуха … Большой медицинский словарь
В данной статье имеется список источников или внешних ссылок, но источники отдельных утверждений остаются неясными из за отсутствия сносок … Википедия
Латеральное коленчатое тело - два клеточных ядра таламуса, расположенные на концах каждого из оптических трактов. К левому телу подходят пути от левой стороны левой и правой сетчатки, к правому соответственно правой стороны сетчатки. Отсюда зрительные пути направляются к… … Энциклопедический словарь по психологии и педагогике
Латеральное коленчатое тело (ЛКТ) - Основной сенсорный центр зрения, расположенный в таламусе участке мозга, играющем по отношению к входящей сенсорной информации роль главного коммутационного устройства. Аксоны, исходящие из ЛКТ, входят в зрительную зону затылочной доли коры … Психология ощущений: глоссарий
ГОЛОВНОЙ МОЗГ - ГОЛОВНОЙ МОЗГ. Содержание: Методы изучения головного мозга..... . . 485 Филогенетическое и онтогенетическое развитие головного мозга............. 489 Bee головного мозга..............502 Анатомия головного мозга Макроскопическое и… … Большая медицинская энциклопедия
Это подкорковый центр, который обеспечивает передачу информации уже в зрительную кору.
У человека эта структура имеет шесть слоёв клеток, как и в зрительной коре. Волокна от сетчатки поступают перекрещенные и неперекрещенные в chiasma opticus. 1-й, 4-й, 6-й слои получают перекрещенные волокна. 2-й, 3-й, 5-й слои получают неперекрещенные.
Вся информация, поступающая к наружному коленчатому телу от сетчатки, упорядочена и сохраняется ретинотопическая проекция. Поскольку волокна входят в наружное коленчатое тело по типу гребёнки, в НКТ нет таких нейронов, которые получают информацию от двух сетчаток одновременно. Из этого следует, что в нейронах НКТ отсутствует бинокулярное взаимодействие. К НКТ поступают волокна от M-клеток и P-клеток. M-путь, сообщающий информацию от крупных клеток, передаёт информацию о движениях объектов и оканчивается в 1-м и 2-м слоях. P-путь связан с цветовой информацией и волокна оканчиваются в 3-м, 4-м, 5-м, 6-м слоях. В 1-м и 2-м слоях НКТ рецептивные поля высокочувствительны к движению и не различают спектральные характеристики (цвет). Такие рецептивные поля в небольшом количестве присутствуют и в других слоях НКТ. В 3-м и 4-м слоях преобладают нейроны с OFF-центром. Это сине-жёлтая или сине-красная + зелёная. В 5-м и 6-м слоях представлены нейроны с ON-центрами в основном красно-зелёные. Рецептивные поля клеток наружного коленчатого тела обладают такими же рецептивными полями, как и ганглиозные клетки.
Отличие этих рецептивных полей от ганглиозных клеток:
1. В размерах рецептивных полей. Клетки наружного коленчатого тела имеют меньшие размеры.
2. У некоторых нейронов НКТ появляется дополнительная тормозная зона, окружающая периферию.
Для клеток с ON-центром такая дополнительная зона будет иметь знак реакции, совпадающий с центром. Эти зоны только у некоторых нейронов, образуются за счёт усиления латерального торможения между нейронами НКТ. Эти слои – основа выживания конкретного вида. У человека – шесть слоёв, у хищников – четыре.
Детекторная теория появилась в конце 1950-х гг. В сетчатке лягушки (в ганглиозных клетках) были обнаружены реакции, которые непосредственно были связаны с поведенческими реакциями. Возбуждение определённых ганглиозных клеток сетчатки приводило к поведенческим реакциям. Этот факт позволил создать концепцию, согласно которой изображение, представленное на сетчатке, обрабатывается специфически настроенными на элементы изображения ганглиозными клетками. Такие ганглиозные клетки имеют специфическое ветвление дендритов, которое соответствует определённой структуре рецептивного поля. Были обнаружены несколько типов таких ганглиозных клеток. В дальнейшем нейроны, обладающие таким свойством, стали называть детекторными. Таким образом, детектор – это нейрон, реагирующий на определённое изображение или его часть. Оказалось, что и у других, более высокоразвитых животных есть возможность выделять специфический символ.
1. Детекторы выпуклого края – клетка активировалась при появлении крупного объекта в поле зрения;
2. Детектор движущегося мелкого контраста – его возбуждение приводило к попытке захвата это объекта; по контрасту соответствует захватываемым объектам; эти реакции связаны с пищевыми реакциями;
3. Детектор затемнения – вызывает оборонительную реакцию (появление крупных врагов).
Эти ганглиозные клетки сетчатки настроены выделять определённые элементы окружающей среды.
Группа исследователей, работавших над этой темой: Летвин, Матурано, Моккало, Питц.
Детекторными свойствами обладают и нейроны других сенсорных систем. Большинство детекторов зрительной системы связано с выделением движения. У нейронов усиливаются реакции при увеличении скорости движения объектов. Детекторы были обнаружены и у птиц, и у млекопитающих. Детекторы других животных непосредственно связаны с окружающим пространством. У птиц были обнаружены детекторы горизонтальной поверхности, что связано с необходимостью приземления на горизонтальные объекты. Также были обнаружены детекторы вертикальных поверхностей, которые обеспечивают собственные движения птиц в сторону этих объектов. Оказалось, что чем выше животное в эволюционной иерархии, тем выше находятся детекторы, т.е. эти нейроны уже могут находиться не только в сетчатке, но и в высших отделах зрительной системы. У высших млекопитающих: у обезьян и человека – детекторы находятся в зрительной коре. Это важно, поскольку специфический способ, который обеспечивает реакции на элементы внешней среды, переносится на вышележащие уровни мозга, и при этом каждому виду животных присущи собственные специфические виды детекторов. В дальнейшем оказалось, что в онтогенезе детекторные свойства сенсорных систем формируются под влиянием окружающей среды. Для демонстрации этого свойства были проделаны эксперименты исследователями, Нобелевскими лауреатами, Хьюбелом и Визелом. Были проделаны эксперименты, доказавшие, что формирование детекторных свойств происходит в самом раннем онтогенезе. Например, использовали три группы котят: одна контрольная и две экспериментальные. Первая экспериментальная была помещена в условия, где в основном присутствовали горизонтально ориентированные линии. Вторая экспериментальная была помещена в условия, где в основном были горизонтальные линии. Исследователи проверяли, какие нейроны сформировались в коре у котят каждой группы. В коре у этих животных оказалось по 50% нейронов, которые активировались и горизонтальными, + 50% вертикальными. Животные, воспитанные в горизонтальной среде, имели в коре значительное количество нейронов, которые активировались горизонтальными объектами, практически не было нейронов, активировавшихся при восприятии вертикальных объектов. Во второй экспериментальной группе была аналогичная ситуация с горизонтальными объектами. У котят обеих горизонтальных групп появились определённые дефекты. Котята горизонтальной среды могли прекрасно прыгать по ступеньками и горизонтальными поверхностям, но плохо проводили движения относительно вертикальных объектов (ножка стола). У котят второй экспериментальной группы была соответствующая ситуация для вертикальных объектов. Данный эксперимент доказал:
1) формирование нейронов в раннем онтогенезе;
2) животное не может адекватно взаимодействовать.
Изменение поведения животных в изменяющейся среде. Каждое поколение имеет свой набор внешних стимулов, которые вырабатывают новый набор нейронов.
Специфические особенности зрительной коры
От клеток наружного коленчатого тела (имеет 6-слойную структуру) аксоны поступают к 4 слоям зрительной коры. Основная масса аксонов наружного коленчатого тела (НКТ) распределяется в четвёртом слое и его подслоях. От четвёртого слоя информация поступает к другим слоям коры. Зрительная кора сохраняет принцип ретинотопической проекции так же, как и НКТ. Вся информация от сетчатки поступает к нейронам зрительной коры. Нейроны зрительной коры, как и нейроны нижележащих уровней, имеют рецептивные поля. Структура рецептивных полей нейронов зрительной коры отличается от рецептивных полей НКТ и клеток сетчатки. Хьюбел и Визел также занимались изучением зрительной коры. Их работа позволила создать классификацию рецептивных полей нейронов зрительной коры (РПНЗрК). Х. и В. Обнаружили, что РПНЗрК имеют не концентрическую, а прямоугольную форму. Они могут быть ориентированы под разными углами, иметь 2 или 3 антагонистических зоны.
Такое рецептивное поле может выделять:
1. изменение освещённости, контраст - такие поля были названы простыми рецептивными полями ;
2. нейроны со сложными рецептивными полями – могут выделять те же самые объекты, что и простые нейроны, но при этом эти объекты могут находиться в любом месте сетчатки;
3. сверхсложные поля - могут выделять объекты, имеющие разрывы, границы или изменение формы объекта, т.е. сверхсложные рецептивные поля могут выделять геометрические формы.
Гештальты – нейроны, выделяющие подобразы.
Клетки зрительной коры могут только формировать некие элементы изображения. Откуда появляется константность, где появляется зрительный образ? Ответ был найден в ассоциативных нейронах, которые также связаны со зрением.
Зрительная система может выделять различные цветовые характеристики. Сочетание оппонентных цветов позволяет выделять различные оттенки. Обязательно участвует латеральное торможение.
Рецептивные поля имеют антогонистические зоны. Нейроны зрительной коры способны возбуждаться периферически на зелёный в то время, как середина возбуждается на действие красного источника. Действие зелёного будет вызывать тормозную реакцию, действие красного будет вызывать возбуждающую реакцию.
Зрительная система воспринимает не только чистые спектральные цвета, но и любые сочетания оттенков. Многие области коры больших полушарий имеют не только горизонтальное, но и вертикальное строение. Это было обнаружено в середине 1970-х гг. Это было показано для соматосенсорной системы. Вертикальная или колончатая организация. Оказалось, что зрительная кора имеет кроме слоёв ещё и вертикально ориентированные колонки. Совершенствование техники регистрации привело к проведению более тонких экспериментов. Нейроны зрительной коры кроме слоёв имеют ещё и горизонтальную организацию. Был проведён микроэлектрод строго перпендикулярно поверхности коры. Все основные зрительные поля в медиальной части затылочной коры. Поскольку рецептивные поля имеют прямоугольную организацию, точки, пятна, любые концентрически объекта не вызывают никакой реакции в коре.
Колонка – вид реакции, соседняя колонка тоже выделяет наклон линии, но от предыдущей он отличается на 7-10 градусов. Дальнейшие исследования показали, что рядом располагаются колонки, у которых угол изменяется с равным шагом. Около 20-22 соседних колонок будут выделять все наклоны от 0 до 180 градусов. Совокупность колонок, способных выделить все градации этого признака, назвали макроколонкой. Это были первые исследования, которые показали, что зрительная кора может выделять не только единичное свойство, но и комплекс – все возможные изменения признака. В дальнейших исследованиях было показано, что рядом с макроколонками, фиксирующими угол, располагаются макроколонки, способные выделять и другие свойства изображения: цвета, направление движения, скорость движения, а также макроколонки, связанные с правой или левой сетчаткой (колонки глазодоминантности). Таким образом, все макроколонки компактно располагаются на поверхности коры. Было предложено совокупности макроколонок называть гиперколонками. Гиперколонки могут анализировать набор признаков изображений, находящихся в локальном участке сетчатки. Гиперколонки – модуль, который выделяет набор признаков в локальном участке сетчатки (1 и 2 идентичные понятия).
Таким образом, зрительная кора состоит из набора модулей, которые анализируют свойства изображений и создают подобразы. Зрительная кора – не конечный этап переработки зрительной информации.
Свойства бинокулярного зрения (стереозрения)
Эти свойства облегчают и животному, и человеку восприятие удалённости объектов и глубины пространства. Для того, чтобы эта способность проявлялась, обязательны движения глаз (конвергентно-дивергентные) на центральную ямку сетчатки. При рассмотрении удалённого объекта происходит разведение (дивергенция) оптических осей и сведение для близко расположенных (конвергенция). Такая система бинокулярного зрения представлена у разных видов животных. Наиболее совершенна эта система у тех животных, у которых глаза располагаются на фронтальной поверхности головы: у многих хищных животных, птиц, приматов, большинство хищных обезьян.
У другой части животных глаза располагаются латерально (копытные, млекопитающие и т.д.). Для них очень важно иметь большой объём восприятия пространства.
Это связано со средой обитания и их местом в пищевой цепочке (хищник - жертва).
При таком способе восприятия пороги восприятия снижаются на 10-15%, т.е. у организмов, обладающих этим свойством, появляется преимущество в точности собственного движений и соотнесении их с движениями цели.
Также существуют монокулярные признаки глубины пространства.
Свойства бинокулярного восприятия:
1. Фузия – слияние полностью идентичных изображений двух сетчаток. При этом объект воспринимается двухмерным, плоскостным.
2. Слияние двух неидентичных изображений сетчаток. При этом объект воспринимается объемно, трехмерно.
3. Соперничество полей зрения. От правой и левой сетчатки поступают два разных изображения. Мозг не может совместить два разных изображения, и поэтому они воспринимаются поочередно.
Остальные точки сетчатки – диспаратные. Степень диспаратности и будет определять, воспринимается ли объект трёхмерно или он будет восприниматься при соперничестве полей зрения. Если диспаратность невелика, то изображение воспринимается трёхмерно. Если диспаратность очень высокая, то объект не воспринимается.
Такие нейроны обнаружены не в 17-м, а в 18-м и 19-м полях.
Чем отличаются рецептивные поля таких клеток: для таких нейронов в зрительной коре рецептивные поля либо простые, либо сложные. В этих нейронах наблюдается различие рецептивных полей от правой и левой сетчатки. Диспаратность рецептивных полей таких нейронов может быть либо вертикальной, либо горизонтальной (см. след. страницу):

Это свойство позволяет лучше адаптироваться.
(+) Зрительная кора не позволяет говорить о том, что в ней формируется зрительный образ, то константность отсутствует во всех областях зрительной коры.
Латеральное коленчатое тело
Латеральное коленчатое тело (наружное коленчатое тело, ЛКТ) - легко распознаваемая структура мозга , которая помещается на нижней латеральной стороне подушки таламуса в виде достаточно большого плоского бугорка. В ЛКТ приматов и человека морфологически определено шесть слоев: 1 и 2 - слои больших клеток (магноцеллюлярные), 3-6 - слои малых клеток (парвоцеллюлярные). Слои 1, 4 и 6 получают афференты от контрлатерального (расположенного в противоположном по отношению к ЛКТ полушарии) глаза, а слои 2, 3 и 5 - от ипсилатерального (расположенного в том же, что и ЛКТ полушарии).
Схематическая диаграмма ЛКТ приматов. Слои 1 и 2 расположены более вентрально, ближе к приходящим волокнам оптического тракта.
Число слоев ЛКТ, участвующих в обработке сигнала, приходящего от ганглиозных клеток сетчаток , различно в зависимости от эксцентриситета сетчатки:
- - при эксцентриситете меньше 1º в обработке участвуют два парвоцеллюлярных слоя;
- - от 1º до 12º (эксцентриситет слепого пятна) - все шесть слоев;
- - от 12º до 50º - четыре слоя;
- - от 50º - два слоя, связанных с контрлатеральным глазом
Бинокулярных нейронов в ЛКТ приматов нет. Они появляются только в первичной зрительной коре.
Литература
- Хьюбел Д. Глаз, мозг, зрение / Д. Хьюбел; Пер. с англ. О. В. Левашова и Г. А. Шараева.- М.: «Мир», 1990.- 239 с.
- Морфология нервной системы: Учеб. пособие / Д. К. Обухов, Н. Г. Андреева, Г. П. Демьяненко и др.; Отв. ред. В. П. Бабминдра. - Л.: Наука, 1985.- 161 с.
- Erwin E. Relationship between laminar topology and retinotopy in the rhesus lateral geniculate nucleus: results from a functional atlas / E. Erwin, F.H. Baker, W.F. Busen et al. // Journal of Comparative Neurology.- 1999.- Vol.407, № 1.- P.92-102.
Wikimedia Foundation . 2010 .
- Абкаик (нефтяное месторождение)
- 75-й полк рейнджеров
Смотреть что такое "Латеральное коленчатое тело" в других словарях:
Латеральное коленчатое тело - два клеточных ядра таламуса, расположенные на концах каждого из оптических трактов. К левому телу подходят пути от левой стороны левой и правой сетчатки, к правому соответственно правой стороны сетчатки. Отсюда зрительные пути направляются к… … Энциклопедический словарь по психологии и педагогике
Латеральное коленчатое тело (ЛКТ) - Основной сенсорный центр зрения, расположенный в таламусе участке мозга, играющем по отношению к входящей сенсорной информации роль главного коммутационного устройства. Аксоны, исходящие из ЛКТ, входят в зрительную зону затылочной доли коры … Психология ощущений: глоссарий
коленчатое тело латеральное - (с. g. laterale, PNA, BNA, JNA) К. т., лежащее на нижней поверхности таламуса латерально от ручки верхнего холмика четверохолмия: место расположения подкоркового центра зрения … Большой медицинский словарь
Зрительная система - Проводящие пути зрительного анализатора 1 Левая половина зрительного поля, 2 Правая половина зрительного поля, 3 Глаз, 4 Сетчатка, 5 Зрительные нервы, 6 Глазодви … Википедия
Структуры мозга - Мозг человека реконструкция на основе МРТ Содержание 1 Мозг 1.1 Prosencephalon (передний мозг) … Википедия
Визуальное восприятие
Зрение - Проводящие пути зрительного анализатора 1 Левая половина зрительного поля, 2 Правая половина зрительного поля, 3 Глаз, 4 Сетчатка, 5 Зрительные нервы, 6 Глазодвигательный нерв, 7 Хиазма, 8 Зрительный тракт, 9 Латеральное коленчатое тело, 10… … Википедия
Зритель - Проводящие пути зрительного анализатора 1 Левая половина зрительного поля, 2 Правая половина зрительного поля, 3 Глаз, 4 Сетчатка, 5 Зрительные нервы, 6 Глазодвигательный нерв, 7 Хиазма, 8 Зрительный тракт, 9 Латеральное коленчатое тело, 10… … Википедия
Зрительная система человека - Проводящие пути зрительного анализатора 1 Левая половина зрительного поля, 2 Правая половина зрительного поля, 3 Глаз, 4 Сетчатка, 5 Зрительные нервы, 6 Глазодвигательный нерв, 7 Хиазма, 8 Зрительный тракт, 9 Латеральное коленчатое тело, 10… … Википедия
Зрительный анализатор - Проводящие пути зрительного анализатора 1 Левая половина зрительного поля, 2 Правая половина зрительного поля, 3 Глаз, 4 Сетчатка, 5 Зрительные нервы, 6 Глазодвигательный нерв, 7 Хиазма, 8 Зрительный тракт, 9 Латеральное коленчатое тело, 10… … Википедия